Гниене на нервните ганглии. ганглии
При гръбначните същества във вегетативната нервна система има три вида синаптично предаване: електрическо, химическо и змишана. Орган с типични електрически синапси е цилиарният ганглий на птиците, който се намира в дълбокия край на основата ябълка на пълен работен ден. Предаването на повикването за събуждане работи практически безпроблемно и в двете посоки. До тихо, което рядко се чува, можете да видите предаването чрез смесени синапси, в които могат да се преценяват едновременно структурите на електрически и химически синапси. Този вид засяга и цилиарния ганглий на птиците. Основният начин за пренасяне на увреждане на вегетативната нервна система е химическият. Vіn zdіysnyuєtsya зад законите за пеене, сред тях виждаме два принципа. Първият (принципът на Дейл) е във факта, че невронът с усма възрастни вижда един медиатор. Както вече стана ясно, редът на основните неврони може да присъства и други предавания участват в техния синтез на речта. Според друг принцип, действието на кожен медиатор върху неврон или ефектор се дължи на естеството на постсинаптичната мембрана на рецептора.
В автономната нервна система има повече от десет вида нервни клетки, които произвеждат като основни различни медиатори: ацетилхолин, норепинефрин, серотонин и други биогенни амини, аминокиселини, АТФ. В зависимост от факта, че основният медиатор се разглежда като крайни аксони на автономните неврони, клетките обикновено се наричат холинергични, адренергични, серотонинергични, пуринергични и др. неврони.
Кожата на медиаторите vikonuє предавателна функция, като правило, в пеещите ланки на дъгата на автономния рефлекс. Така ацетилхолинът се вижда в окончанията на всички преганглионни симпатикови и парасимпатикови неврони, както и в повечето постганглионни парасимпатикови окончания. В допълнение, част от постганглионните симпатикови влакна, които инервират потните жлези и, може би, вазодилататори на скелетни язви, също насърчават прехвърлянето на ацетилхолин за помощ. От своя страна норепинефринът е медиатор в постганглионните симпатикови окончания (заради нервите на потните кухини и симпатиковите вазодилататори) - съдът на сърцето, черния дроб, далака.
Медиатор, който работи в пресинаптичните терминали под притока на нервни импулси, който идва, взаимодействайки със специфичен рецепторен протеин на постсинаптичната мембрана и създавайки сложна връзка с него. Протеинът, взаимозаменяемо ацетилхолин, се нарича холинергичен рецептор, адреналин или норадреналин - адренорецептор и т.н. Мястото на локализация на рецепторите на различни медиатори е не само постсинаптична мембрана. Установено е, че са разкрити специални пресинаптични рецептори, които участват в механизма на индуциране на регулация на медиаторния процес в синапса.
Крим холино-, адрено-, пуринорецептори, в периферната част на вегетативната нервна система е рецептори на пептиди, допамин, простагландини. Всички видове рецептори, които са открити в периферната част на вегетативната нервна система, са открити в пре-постсинаптичните мембрани на ядрените структури на ЦНС.
Характерната реакция на вегетативната нервна система е рязко повишаване на чувствителността към медиатори след денервация на органи. Например, след ваготомия, органът може да бъде чувствителен към ацетилхолин и, очевидно, след симпатектомия, към норепинефрин. Трябва да се има предвид, че в основата на това явление е рязкото увеличаване на броя на рецепторите в постсинаптичната мембрана, както и намаляване на активността на ензимите, които разцепват медиатора (ацетилхолин-естераза, моноаминооксидаза). и др.).
В автономната нервна система, най-изявените ефекторни неврони, има и специални клетки, които имитират постганглионни структури и определят тяхната функция. Пренасянето на възбуда към тях се извършва по специален химичен начин, а вонята по ендокринен. Qi клетките отнеха името на преобразувателите. Аксоните им не образуват синаптични контакти с ефекторни органи, а завършват близо до съдовете, с които образуват т. нар. хемални органи. Преди трансдукторите, напредващият клитиний може да се припише: 1) хромофинен клитиния на медуларната топка на наднервните гънки, като холинергичен трансмитер на преганглионния симпатиков край, показателен за адреналин и норепинефрин; 2) юкста-гломерулни клетки на нирк, як, което показва адренергично предаване на постганглионното симпатиково влакно към зрението на кръвния поток на ренина; 3) неврони на хипоталамичните супраоптични и паравентрикуларни ядра, които реагират на синаптичния приток от различно естество на вазопресин и окситоцин; 4) неврони на ядрата на хипоталамуса.
За помощ може да се създаде ролята на главните класически медиатори фармакологични препарати. Например, никотинът предизвиква ефект, подобен на този на ацетилхолина, когато се прилага върху постсинаптичната мембрана на постганглионния неврон, докато в същото време етер холинът и токсинът от мухоморка мускарин се сгъват към постсинаптичната мембрана на ефективната клетка на висцералната орган. Също така, никотинът участва в интерневронното предаване във вегетативния ганглий, мускаринът - в невро-ефекторното предаване във висцералния орган. На тази основа е важно, че има два вида холинергични рецептори: никотинови (N-холинергични рецептори) и мускаринови (М-холинергични рецептори). В зависимост от чувствителността към различни катехоламини, адренорецепторите се подразделят на α-адренергични рецептори и β-адренергични рецептори. Тази причина е установена с помощта на фармакологични препарати, които са вибрационни за пеещия тип адренорецептори.
В редица висцерални органи, които реагират на катехоламини, адренорецепторите могат да бъдат обидени, но резултатите от тяхното пробуждане като правило са протоиленги. Например, в кръвоносните съдове на скелетните мембрани има α- и β-адренергични рецептори. Увреждането на α-адренергичните рецептори води до звънене, а β-адренергичните рецептори до разширяване на артериолите. И двата вида адренергични рецептори и прояви в чревния тракт, протео-реакция на органа в случай на пробуждане на кожата недвусмислено се характеризират с галванична активност на гладки злокачествени клетки. В сърцето и бронхите няма α-адренергични рецептори и медиатор на взаимодействие само с β-адренергични рецептори, което е придружено от ускоряване на сърдечната честота и разширяване на бронхите. В същото време норепинефринът предизвиква най-голямо стимулиране на β-адренергичните рецептори в сърцето, а слабата реакция на бронхите, трахеята, съдовете и първите започва да се нарича β1-адренергични рецептори, други - β2-адренергични рецептори.
Когато атакуват мембраната на гладката мукозна клетка, епинефринът и норепинефринът активират аденилциклазата, която присъства в клетъчната мембрана. За наличието на Mg2+ йони, този ензим катализира превръщането на cAMP (цикличен 3,5"-аденозин монофосфат) в АТФ в клетките. Останалата част от продукта, сама по себе си, изисква редица физиологични ефекти, активиращи енергийния обмен, стимулиране на сърдечната дейност.
Особеностите на адренергичния неврон са тези, които могат да надлингвистично дълги тънки аксони, които се образуват в органите и образуват плътни плексуси. Общата дължина на такива аксонални терминали може да достигне 30 см. В хода на терминалите има числени разширения - разширени вени, в които се синтезира, съхранява и се вижда като медиатор. С пристигането на импулса, норепинефринът веднага се вижда от численото разширение, разпръсквайки голямата площ на гладката тъкан на езика. По този начин деполяризацията на m'yazovykh клитинум е придружена от едночасов недостиг на целия орган.
Rіznі медицински грижи, който се прилага върху ефекторния орган, подобно на този на постганглионното влакно (симпатикова, парасимпатикова кльощава), отне името на миметиците (адрено-, холиномиметици). Инструкции от речта и речта, вибриращи блокиращи функцията на рецепторите в постсинаптичната мембрана. Наричат се блокери на ганглии. Например, амониевите вибрират вибриращи Н-холинергичните рецептори, а атропинът и скополаминът - М-холинергичните рецептори.
Класическите медиатори печелят не само функцията за предаване на възбуда, но и жизнеността и жизнената биологична активност. Сърдечно-съдовата система е най-чувствителна към ацетилхолина, той стимулира и засилва подвижността на билковия тракт, активирайки дейността на билковите заболявания наведнъж, скъсява мускулите на бронхите и намалява бронхиалната секреция. При инфузията на норепинефрин се наблюдава повишаване на систоличното и диастолното налягане без промяна на сърдечния ритъм, сърдечните контракции се увеличават, секрецията на тръбата и червата намалява, гладката мускулатура на червата се отпуска. г. Адреналинът се характеризира с по-голямо разнообразие от диапазони. За допълнителна едночасова стимулация ино-, хронодромотропните функции на адреналина повишават сърдечната честота. Адреналинът повишава и спазмолитичния ефект върху бронхиалните мускули, ревматоидната подвижност на билковия тракт, отслабва стените на органите, а също така разклаща сфинктерите, секрецията на билковия тракт.
Серотонин (5-хидрокситриптамин) е открит в тъканите на всички животински видове. В мозъка вените са по-важни в структурите, които регулират регулирането на висцералните функции, на периферията те се произвеждат от ентерохромафинни клитини на червата. Серотонинът е един от основните медиатори на метасимпатиковата част на автономната нервна система, който играе основна роля в предаването на неврорефлектора, а също така функционира като медиатор в централните камери. Има три вида серотонинергични рецептори - D, M, T. D-тип рецептори са локализирани главно в гладки меса и са блокирани от диетиламида на лизергиновата киселина. Взаимодействието на серотонина с тези рецептори е придружено от лека заболеваемост. Рецепторите от М-тип се характеризират с по-големи автономни ганглии; блокиран от морфин. Свързвайки се с cimi рецептори, предавайки пулсиращ ганглиостимулиращ ефект. Рецепторите от Т-тип, открити в рефлексогенните зони на сърцето и краката, се блокират от тиопендол. Поради наличието на ци рецептори, серотонинът участва в развитието на коронарни и белодробни хеморефлекси. Изграждането на серотонин оказва директен натиск върху гладките мускули. В съдовата система се проявява в констрикторни и дилататорни реакции. При директно дишане мускулите на бронхите се съкращават, с рефлекторно действие се променят дикалният ритъм и легеновата вентилация. Билковата система е особено чувствителна към серотонин. При въвеждането на серотонин той реагира с настъпваща спастична реакция, която преминава в ритмична бързина с повишен тонус и галванична активност, която завършва.
За богатите висцерални органи, характерната пуринергична трансмисия е кръстена на факта, че аденозин и инозин, продукти на разграждане на пурин, се наблюдават по време на стимулация на пресинаптичните терминали. Медиатор в тази посока е пресинаптичният терминал на ефекторните неврони в метасимпатиковата част на вегетативната нервна система.
АТФ, който се наблюдава в синаптичната празнина, взаимодейства с пуринорецептори на постсинаптичната мембрана от два типа. Пуринорецепторите от първия тип са по-чувствителни към аденозин, другият - към АТФ. Действието на медиатора е директно насочено към гладката мускулатура, което се проявява в появата на релаксация. В механизма на чревното задвижване пуринергичните неврони преминават от антагонистичната галванична система на главата към възбуждащата холинергична система. Пуринергичните неврони участват в намалената ниска галванизация, в механизма на рецептивния релаксин, отпускането на стравохида и аналния сфинктер. Недостигът на червата, който се обвинява за пуринергично викане за релаксация, осигурява правилното преминаване на хранителните гърди.
Сред медиаторите можете да използвате хистамин. Vіn широки разширения в различни органи и тъкани, особено в билковия тракт, legenia, shkіrі. Сред структурите на вегетативната нервна система най-голямо количество хистамин се намира в постганглионните симпатикови влакна. На фона на реакциите в определени тъкани са открити специфични хистаминови (H-рецептори) рецептори: H1- и H2-рецептори. Класическо действие на хистамин и повишено капилярно проникване и недостиг на гладка мускулатура. В свободно състояние хистаминът намалява кръвното налягане, променя сърдечната честота, стимулира симпатиковия ганглий.
За интерневронното предаване на възбуждане в ганглиите на вегетативната нервна система може да се използва галваничното инжектиране на GABA. Като посредник можете да вземете съдбата на виновната пресинаптична галванизация.
Големи концентрации на различни пептиди, особено вещество P, в тъканите на билковия тракт, хипоталамуса, задната кора на гръбначния мозък, както и ефектите от стимулиране на останалите и други индикации, станаха основата за употребата на вещество P като медиатор на чувствителните нервни клетки.
При класическите медиатори и "кандидатите" сред медиаторите регулирането на дейността на викарните органи взема участие в участието на голям брой биологично активни речи - мускулните хормони. Вонята регулира тонуса, коригира притока върху дейността на вегетативната нервна система, има важна роля в координацията на неврохуморалната трансмисия, в механизмите на зрението на тези медиатори.
В комплекса от активни фактори пространството заемат простагландините, които са богато разположени във влакната на изпъкналия нерв. Звуци на смрад се виждат спонтанно при изблик на стимулация. Основният клас простагландини: E, G, A, B. Основният ефект е стимулиране на гладките лигавици, потискане на секрецията на лигавицата, отпускане на мускулите на бронхите. Върху сърдечно-съдовата система вонята може да бъде различно насочена: простагландините от клас А и Е наричат вазодилатация и хипотония, клас G - вазоконстрикция и хипертония.
Синапсът на ANS е като сън, като централен. Показано е значително разширяване на хеморецепторите в постсинаптичните мембрани. Предаването на нервните импулси от преганглионните влакна към невроните на всички автономни ганглии се медиира от Н-холинергични синапси, tobto. синапси на постсинаптичната мембрана на такива ротации на никотин-чувствителни холинергични рецептори. Постганглионните холинергични влакна работят върху клитините на висцералните органи (сол, SMC на органите на ецване, тънки съдове) М-холинергични синапси. Това е постсинаптична мембрана, която се съпротивлява на мускариновите рецептори (блокер на атропина). В тези други синапси предаването на възбудата се медиира от ацетилхолин. М-холинергичните синапси могат да имат стимулираща инфузия върху гладката слуз на билковия канал, сиеховивидната система (сфинктерния сфинктер), вените на SCT. Все пак смрадите променят бдителността, проводимостта, недостига на сърдечно месо и вика от отпускането на някои съдове на главата и таза.
Постганглионните симпатикови влакна установяват 2 вида адренергични синапси на ефекторите – а-адренергични и b-адренергични. Постсинаптичната мембрана на първия a1-i a2 е адренорецептори. При NA на a1-адренергичните рецептори има сондиране на артерии и артериоли вътрешни органии shkіri, задух на лигавиците на матката, сфинктери на лигавиците-чревния тракт, и в същото време отпускане на други гладки лигавици на билковия канал. Постсинаптичните b-адренергични рецептори също се подразделят на b1 - и b2 - връхчета. b1-адренергични рецептори, roztashovani в клитини на сърдечно месо. Когато са засегнати, се наблюдават бдителността, проводимостта и късостта на кардиомиоцитите. Активирането на b2-адренергичните рецептори води до разширяване на съдовете на краката, сърцето и скелетните язви, отпускане на гладки бронхиални язви, sich michur, галванизиране на подвижността на органите ецване
Освен това бяха разкрити постганглионните влакна, които работят върху клитините на вътрешните органи в хистаминергичните, серотонинергичните, пуринергичните (АТФ) синапси.
2. Становище на Павлов за 1 и 2 сигнални системи.
Сигналната система е система от умствени и безумни рефлексни връзки на висшата нервна система на съществата (включително хората) и излишния свят. Rozrіznyayut pershu и други сигнални системи.
Терминът е одобрен от акад. И. П. Павлов.
Първата сигнална система на практика се намира във всички същества, въпреки че другата система присъства само при хората и, може би, в някои подобни на кит. Tse pov'yazano z tim, scho само хората zdatna оформят абстрактен образ на ситуацията. След като запомни думата „лимон“, човек може да покаже кое вино е кисело и как да се намръщи, ако го чуете, след това запомнянето на думата извиква образ в паметта (използва се другата сигнална система); yakscho, с когото започна движението на линията, роботът на първата сигнална система.
Предмет на изучаване на физиологията е висшата нервна дейност.
Друга сигнална система е специален вид по-голяма нервна дейност на човек, системата от "сигнални сигнали", която върви по посока на дивата (макар и не същата) със съществата от първата сигнална система - очевидно е , се проявява, че лежи до точката на светлината. Да вървим, като различна сигнална система, като семиотична система от значения – че „какво да отидем в кората на вида на нашите органи е други сигнали, сигнали от сигнали. Вонята е двустранен акт и е позволено да се влоши, иска да стане наша специална личност, особено човешка, по-скоро ум, който създава гръбнака на човешкия емпиризъм, а, нареши, и науката - за най-важния ориен - т на човешки същества с наука. аз П. Павлов (1932).
Мозъкът на съществото се показва само посред нощ, следват звуци и други рими; Очевидно те обвиняват, за да установят първата сигнална система за дейност.
В процеса на еволюция на света на създанията на етапа на развитие на кочана на вида Homo sapiens се е появила своеобразна сигнална система, която осигурява активното и колективно адаптивно обичайно поведение, което създава разнообразни, приети за групата дума на сигналната система: П. Павлова, се превръща в „сигнал на сигналите”. Възможност за друга система за сигнализация - Viknnynennya Movy iov, сигнални системи People's rhodiers, de Umovnі (Dovіlnі) Сигнални сигнали Іndivіda Nabuvyuti Sovnі, Princeyі Group Valida Ta Snowstick, подмладена на знаците на Movy в директната Rosuminіvіv Túsívív níssívіív nіssіvіvіyyiy Животът на рода Homo, който се предава чрез движение от поколение на поколение.
При ул. Вивчени с. с. Гръбнакът беше по-важен от натрупването на факти, които характеризират значението на основните функции на словесните сигнали, че buv - разширяването на нервните механизми на словото. Установено е, че развитието на думата се развива в резултат на развитието на система от мисловни връзки (разв. Интелигентно отражение); собственото му значение е като количество обаждания, а характерът му е: повикване, вибрации в часа на активността на детето, което улеснява процеса на обостряне. Когато има вербални сигнали, има постоянни промени в бдителността, голяма сила, честота и тривалност на електрическите разряди в нервните клетки на пеещите пункции на мозъка. Росвиток ул. с. с. - резултат от дейността на всички морбили на големия пивкул; невъзможно е да се наруши този процес от функцията на такъв нечист мозък. Dosledzhennyah V.S.S. в лабораторията по висша невродинамика и психология на висшите когнитивни процеси. аз Бойка показа валидността на сватбата I. П. Павлова за динамични тимча връзки V.s.s. При развитието на идеите И. П. Павлова и Е. А. Бойка в училище Й. А. естеството на връзката на промоцията с мисъл, промоцията със специалната промоция; СПЕЦИАЛНО РАЗПРЕДЕЛЕНИЕ НА DIYTYACHO MOVA TA II ИМА ROSERBLENIY NOVIE МЕТОДИ AN ANALIZA OF PUBLICHNYY VOTTERIV (Интент-аниназЗ), ШОО тозволёє Стойтую мирома морова морова и други, at the led простовохромова сила и др. .
Іstotnim резерва за гостите, след doslіdzhen zalishayutsya Проблеми tipologії kolosalnye іndivіdualnih vіdmіnnostey в vzaєmozv'yazkah zagalnogo че spetsіalnogo tipіv БНД неокортекса, че emotsіyno-volovoї че mimovіlnoї regulyatsії dіyalnostі че spіlkuvannya, боде Scho як слабо представени в fіzіologії БНД, така че аз в psiholіngvіstichnih doslіdzhennyah че в antropologіchnіy линии.
Подробности
ганглиибъдете себе си клъстер от мултиполярни (един аксон и цаца дендрити) неврони(Vid kіlkoh kitin до десетки хиляди). Екстраорганичните (симпатиковите) ганглии могат да бъдат добре показана добра тъканна капсула, като продължение на периневриума. Парасимпатиковите ганглии чуруликат, звучащи, при интрамурални нервни клюки. Ганглиите на интрамуралните плексуси, подобно на други вегетативни възли, отмъщават на вегетативни неврони на рефлекторни дъги. Мултиполярните неврони с диаметър 20-35 µm са дифузно разпространени, кожните неврони се дренират от ганглийни глиоцити.
Освен това описано невроендокринни, хеморецепторни, биполярни и в някои гръбначни и униполярни неврони. В симпатичните ганглии има интензивно флуоресцентни клетки (MIF-клетки) с къси израстъци и голям брой гранулирани луковици в близост до цитоплазмата. Вонята вижда катехоламини и се включва в предаването на импулси от преганглионните нервни влакна към симпатиков еферентен неврон. Клетките се наричат интерневрони.
Сред великите мултиполярни невронивегетативен ганглий razrіznyayut: ruhovі (Clintini Dogel 1-ви тип), чувствителен (Clintini Dogel II тип) и асоциативен (Clіtini Dogel III тип). Моторните неврони развиват къси дендрити с ламеларни разширения („рецептивни майдани“). Аксонът на тези клетки е дълъг, простира се отвъд интерганглия в склада на тънки постганглионни немиелинизирани нервни влакна и завършва на гладки миоцити на вътрешните органи. Клитините от 1-ви тип се наричат преаксонални неврони. Неврони от II-ти тип - равни nevnovidrostkovy нервни клетки. В горната част на тялото има 2-4 израстъци, а аксонът е важен в средата. Не razgaluzhuyuchis, тийнейджърите отиват далеч в тялото на неврона. Техните дендрити могат да бъдат чувствителни към нервните окончания и аксонът ще завършва върху телата на невроните на движещите се неврони в съдовите ганглии. Клетките тип II са чувствителни неврони на мускулните рефлекторни вегетативни дъги. Догеловите клетки от III тип са подобни по форма на вегетативните неврони от II тип, тези дендрити не се простират между ганглия, а невритът е направо до малките ганглии. Много известни хора са насърчавани от различни видове чувствителни неврони.
По този начин в периферните вегетативни ганглии има макулни рефлексни дъги, които са съставени от чувствителни, груби и евентуално асоциативни вегетативни неврони.
Интрамуралните вегетативни ганглии в стените на билковия тракт се възкресяват от същите, в техния склад, редица рушови холинергични неврони и галванични неврони. Вонята е представена от адренергични и пуринергични нервни клетки. Останалият медиатор е пуринов нуклеотид. В интрамуралните вегетативни ганглии също растат пептидергични неврони, които виждат вазоинтестиналния пептид, соматостатин и редица други пептиди, с помощта на които се засяга невроендокринната регулация и модулирането на дейността на тъканите и органите в билковата система.
ацетилхолин- никотинови (кураре блок, хексаметоний), мускаринови (атропинов блок) рецептори. Активиране на рецептора → генериране на EPSP. Shvidky EPSP (N-cholinocer)→отваряне на йонни канали. Повишена EPSP (M-холинергична)→prignichennya M-струм, zumovlenogo напредък на K-проводимост.
Невропептиди- Работете като невромодулатори.

Енкефалини, реч П, лулиберин, невротензин, соматостатин - симп. ганглии (+Ach)
Катехоламини(NA, допамин) – невротрансмитери на други клетки поради интензивна флуоресценция.
Невропептид Y, соматостатин - симптом. постганглионарна.
Симпатикова постганглионика: NA, ATP, невропептид U.
α1→инозотолтрифосфат, диацилглицерол. α2→G-протеин активиране, ↓cAMP.
β→G-протеин→AC→cAMP
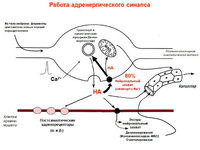
Винятки: медиатор Ach, мускаринови рецептори.
Парасимп. постганглионарни: Ach, VIP, NO, соматостатин, ATP, опиоидни пептиди.
M1 (висока споридност към пирензепин) - повишена секреция на киселина от клитините на шлаката, M2 (ниска) - повдига сърцето. ритъм, отделяне на слуз и шлаки.
Ризноманитна дия:
- Конкретно второ. посредници: M2 може да действа IP3, и може да in-gt AC, променяйки cAMP.
-Дия по Kta Sa-канали
- NO → гуанилатциклаза → cGMP → cGMP-изчерпана протеин киназа → отслабеното освобождаване на лигавицата се абсорбира върху ендотела.
В автономната нервна системаотделят централните и периферните клонове. Централните клонове на симпатиковата нервна система са представени от ядрата на рогата на прасеца на тораколумбалния клон на гръбначния мозък. В парасимпатиковата нервна система централните нерви включват ядрата на средния мозък и ластовича опашка, както и ядрата на рогата на бицепса на сакралния гръбначен мозък. Парасимпатиковите влакна на краниобулбарната камера излизат от складове III-ї, VII-ї, IX-ї и Х-двойкачерепни нерви.
Периферни признаци на вегетативната нервна системаизпълнени с нервни стовбури, ганглии и клюки.
Вегетативни рефлекторни дъгите започват с чувствителен неврон, който лежи близо до гръбначния възел (ганглий), като в соматичния рефлексен възел. Асоциативни неврони perebuvayut в рогата на гръбначния мозък. Тук нервните импулси се предават на проксималните преганглионни неврони, чиито юноши лишават централното ядро и достигат до автономните ганглии, където предават импулсите на грубия неврон. В кръстовището с cym нервните влакна на преганглионните и постганглионните са разделени. Първият от тях лишава централната нервна система в склада на вентралните корени на гръбначните нерви и черепните нерви. Както в симпатиковата, така и в парасимпатиковата системи, преганглионните нервни влакна се припокриват с холинергичните неврони. Аксоните на невроните, разпространени във вегетативните ганглии, се наричат постганглионарни. Вонята не осъществява директен контакт с ефекторните клетки. Тези терминали образуват разширения - разширени вени със собствено темпо, в чийто склад се докупуват луковиците на посредника. В областта на разширените вени няма глиална туника и невротрансмитер, наблюдаван в средата на хранопровода, се влива в ефекторните клетки (например върху клитините, гладките миоцити и др.).
в периферните ганглиисимпатиковата нервна система, като правило, има адренергични еферентни неврони (за малък неврон, който може да синаптични връзки с потни отлагания, десимпатиковите неврони са холинергични). В парасимпатиковите ганглии аферентните неврони обикновено са холинергични.
ганглиие натрупване на мултиполярни неврони (брой клетки до десетки хиляди). Екстраорганичните (симпатиковите) ганглии могат да бъдат добре показана добра тъканна капсула, като продължение на периневриума. Парасимпатиковите ганглии чуруликат, звучащи, при интрамурални нервни клюки. Ганглиите на интрамуралните плексуси, подобно на други вегетативни възли, отмъщават на вегетативни неврони на рефлекторни дъги. Мултиполярните неврони с диаметър 20-35 µm са дифузно разпространени, кожните неврони се дренират от ганглийни глиоцити. Освен това са описани невроендокринни, хеморецепторни, биполярни и в някои гръбначни и униполярни неврони. В симпатичните ганглии има интензивно флуоресцентни клетки (MIF-клетки) с къси израстъци и голям брой гранулирани луковици в близост до цитоплазмата. Вонята вижда катехоламини и се включва в предаването на импулси от преганглионните нервни влакна към симпатиков еферентен неврон. Клетките се наричат интерневрони.
Сред големите мултиполярни неврониВегетативните ганглии се делят на: рухови (клетки на Догел от I-ти тип), чувствителни (клетки на Dogel от II-ти тип) и асоциативни (клетки на Dogel от III-ти тип). Моторните неврони развиват къси дендрити с ламеларни разширения („рецептивни майдани“). Аксонът на тези клетки е дълъг, простира се отвъд интерганглия в склада на тънки постганглионни немиелинизирани нервни влакна и завършва на гладки миоцити на вътрешните органи. Клитини тип I се наричат преаксонални неврони. Тип II неврони - rіvnovidrostkovі нервни клетки. В горната част на тялото има 2-4 израстъци, а аксонът е важен в средата. Не razgaluzhuyuchis, тийнейджърите отиват далеч в тялото на неврона. Техните дендрити могат да бъдат чувствителни към нервните окончания и аксонът ще завършва върху телата на невроните на движещите се неврони в съдовите ганглии. Clitini тип II - чувствителни неврони на мускулните вегетативни рефлекторни дъги. Клетките на Догел тип III са подобни по форма на тялото на вегетативни неврони от тип II, тези дендрити не се простират между ганглиите и невритът е направо до малките ганглии. Много известни хора са насърчавани от различни видове чувствителни неврони.
По такъв начин, периферни автономни ганглиие mіstsevі рефлексни дъги, като чувствителни, rukhovih и, вероятно, асоциативни вегетативни неврони.
Интрамурални автономни ганглиина станцията на билковия тракт има такива, които са в склада, обвивка на холинергични неврони, е galmіvnі неврони. Вонята е представена от адренергични и пуринергични нервни клетки. Останалият медиатор е пуринов нуклеотид. В интрамуралните вегетативни ганглии също растат пептидергични неврони, които виждат вазоинтестиналния пептид, соматостатин и редица други пептиди, с помощта на които се засяга невроендокринната регулация и модулирането на дейността на тъканите и органите в билковата система.
Основна видео анатомия на вегетативната нервна система (ANS)
В случай на проблеми от един поглед, изтеглете видеото отстраниAvtonomnі (vegetativnі) nervovі vuzli (ganglії) mozhut roztashovuvatisya хребет vzdovzh (paravertebralnі ganglії) abo popered Demba (prevertebralnі ganglії) и takozh in stіntsі organіy: sericite, sericite, sericite, sericite, bronhіov, severtebralnі ganglіy, severtebralnі ganglіy. Понякога смрадта може да изглежда като други (от декилко клитин до декилкох десетки клитин) клъстери от неврони, разпространяващи се по хода на някои нерви или интрамурално (микроганглии). Преганглионните влакна (миелин) се изкачват до вегетативните възли, за да отмъстят за израстъците на клетките, чиито тела лежат близо до централната нервна система. Ци влакната силно се огъват и установяват числено синаптично завършване на клитините на вегетативните възли. Причината за това е конвергенцията на голям брой терминали от преганглионни влакна върху кожния неврон на ганглия. Във връзка с наличието на синаптична трансмисия, вегетативните възли се довеждат до нервните центрове от ядрен тип.
Вегетативните нервни възли според функционалния признак и локализация се подразделят на:
красива;
парасимпатиков.
Симпатични нервни възли(para-prevertebral) отрязват преганглионните влакна от клитина, разпределени във вегетативните ядра на гръдния и напречния сегмент на гръбначния мозък. Невротрансмитерът на преганглионните влакна е ацетилхолинът, а на постганглионните - норепинефрин (заради потната пот и други кръвоносни съдове, които могат да имат симпатикова холинергична инервация). В нодулите се появяват невротрансмитери на Krym tsikh, енкефалини, реч P, соматостатин, холецистокинин.
Парасимпатикови нервни възли(интрамурални, които лежат близо до органите или възлите на главата) отрязват преганглионните влакна от клитина, разпределени във вегетативните ядра на дугласа и средния мозък, както и черепния мозък на гръбначния мозък. Ци влакната изпълват централната нервна система в складове 3, 7, 9, 10 двойки черепни нерви и предните коринти на черепните сегменти на гръбначния мозък. Невротрансмитерът на пре-постганглионните влакна е ацетилхолинът. Ролята на медиатори в тези ганглии се играе от серотонин, АТФ и евентуално пептиди.
Повечето от вътрешните органи могат да имат подкожна вегетативна инервация, така че постганглионните влакна да могат да бъдат отстранени от клитините, разпространявани като в симпатиковите и парасимпатиковите възли. Реакциите, опосредствани от клитини на симпатиковите и парасимпатиковите възли, често причиняват продължително изправяне, например: симпатиковата стимулация е силна, а парасимпатиковата - засилва сърдечната дейност.
Централният план на съществуването на симпатиковите и парасимпатиковите нервни възли е подобен. Вегетативният вузол е покрит с тъканна капсула на далака и дифузно или с групи от гниене на тялото от многополярни неврони, техните израстъци в очевидно немиелинизирани или по-вероятно миелинизирани влакна и ендоневрий. Тялото на невроните може да има неправилна форма, ексцентрично разпространение на ядрото, изостряне (звучи не точно) черупки от глиални клитин-сателити (мантийни глиоцити). Често има богати ядрени и полиплоидни неврони.
Интрамуралните връзки и връзките с тях, водещи пътища чрез тяхната висока автономност, сгъваема организация и специфичност на медиаторния обмен, се виждат от някои автори в самостоятелен метасимпатичен клон на вегетативната нервна система. Zokrema, общият брой на невроните в интрамуралните възли на червата е по-висок, по-нисък в близост до гръбначния мозък и поради сгъването на тяхната взаимозависимост в регулацията на перисталтиката и секрецията, те са свързани с миникомпютъра.
Три типа неврони са описани в интрамуралните възли:
Аферентните неврони на дългия аксон (клетки тип I на Догел) са числено по-добри. Това са големи или средни разширения на аферентните неврони с къси дендрити и дълъг аксон, който е право отвъд границата на работния орган, върху клитините на чиито вени установява рушови или секреторни окончания;
Rivnovidrostchasti аферентни неврони (тип II clitiny на Dogel) за отмъщение на дълги дендрити и аксони, които излизат извън междинния ганглий в шева и установяват синапси на clitiny I и III тип. Ци клитини, може би, влизат като рецепторна ланка в склада на рефлексните дъги на мачтата, яки се затварят без нервен импулс, навлизащ в централната нервна система. Наличието на такива дъги се потвърждава от запазването на функционално активни аферентни, асоциативни и аферентни неврони в трансплантирани органи (например сърцето);
асоциативни клитини (клитини на Догел от III тип) са мъжките инсерционни неврони, които пораждат цаца от тип I и II клитини, морфологично подобни на клитините на Dogel от тип II. Дендритите на тези клитини не излизат извън междувъзлията, но аксоните се изправят към други възли, установявайки синапси върху клитини тип I.