Нервові ганглії розташування. Ганглії
У хребетних тварин в автономній нервовій системі є три види синаптичної передачі: електрична, хімічна та змішана. Органом із типовими електричними синапсами є циліарний ганглій птахів, що лежить у глибині очниці біля основи очного яблука. Передача збудження здійснюється практично без затримки в обох напрямках. До тих, що рідко зустрічаються, можна віднести і передачу через змішані синапси, в яких одночасно сусідять структури електричних і хімічних синапсів. Цей вид також уражає циліарного ганглія птахів. Основним способом передачі порушення в автономній нервовій системі є хімічний. Він здійснюється за певними закономірностями, серед яких виділяють два принципи. Перший (принцип Дейла) у тому, що нейрон з усіма відростками виділяє один медіатор. Як стало тепер відомо, поряд з основним у цьому нейроні можуть бути присутніми також інші передавачі та речовини, що беруть участь у їх синтезі. Згідно з другим принципом, дія кожного медіатора на нейрон або ефектор залежить від природи постсинаптичної мембрани рецептора.
В автономній нервовій системі налічують більше десяти видів нервових клітин, які продукують як основні різні медіатори: ацетилхолін, норадреналін, серотонін та інші біогенні аміни, амінокислоти, АТФ. Залежно від того, який основний медіатор виділяється закінченнями аксонів автономних нейронів, ці клітини прийнято називати холінергічними, адренергічними, серотоіінергічними, пуринергічними і т. д. нейронами.
Кожен із медіаторів виконує передатну функцію, як правило, у певних ланках дуги автономного рефлексу. Так, ацетилхолін виділяється в закінченнях всіх прегангліонарних симпатичних та парасимпатичних нейронів, а також більшості постгангліонарних парасимпатичних закінчень. Крім того, частина постгангліонарних симпатичних волокон, що іннервують потові залози і, мабуть, вазодилататори скелетних м'язів, також здійснюють передачу за допомогою ацетилхоліну. У свою чергу норадреналін є медіатором у постгангліонарних симпатичних закінченнях (за винятком нервів потових залоз та симпатичних вазодилататорів) – судин серця, печінки, селезінки.
Медіатор, що звільняється в пресинаптичних терміналах під впливом нервових імпульсів, що приходять, взаємодіє зі специфічним білком-рецептором постсинаптичної мембрани і утворює з ним комплексне з'єднання. Білок, з яким взаємодіє ацетилхолін, зветься холінорецептором, адреналін або норадреналін - адренорецептором і т. д. Місцем локалізації рецепторів різних медіаторів є не тільки постсинаптична мембрана. Виявлено існування та спеціальних пресинаптичних рецепторів, які беруть участь у механізмі зворотного зв'язку регуляції медіаторного процесу у синапсі.
Крім холіно-, адрено-, пуринорецепторів, в периферичній частині автономної нервової системи є рецептори пептидів, дофаміну, простагландинів. Всі види рецепторів, спочатку виявлені в периферичній частині автономної нервової системи, були знайдені потім в пре-і постсинаптичних мембран ядерних структур ЦНС.
Характерною реакцією автономної нервової системи є різке підвищення її чутливості до медіаторів після денервації органів. Наприклад, після ваготомії орган має підвищену чутливість до ацетилхоліну, відповідно після симпатектомії - до норадреналіну. Вважають, що в основі цього явища лежить різке зростання числа відповідних рецепторів постсинаптичної мембрани, а також зниження вмісту або активності ферментів, що розщеплюють медіатор (ацетилхолін-естераза, моноаміноксидаза та ін.).
В автономній нервовій системі, крім звичайних ефекторних нейронів, існують ще спеціальні клітини, що відповідають постгангліонарним структурам і виконують їх функцію. Передача збудження до них здійснюється звичайним хімічним шляхом, а вони відповідають ендокринним способом. Ці клітини отримали назву трансдукторів. Їхні аксони не формують синаптичних контактів з ефекторними органами, а вільно закінчуються навколо судин, з якими утворюють так звані гемальні органи. До трансдукторів відносять такі клітини: 1) хромафінні клітини мозкового шару надниркових залоз, які на холінергічний передавач прегангліонарного симпатичного закінчення відповідають виділенням адреналіну та норадреналіну; 2) юкста-гломерулярні клітини нирки, що відповідають на адренергічний передавач постгангліонарного симпатичного волокна виділенням у кров'яне русло реніну; 3) нейрони гіпоталамічних супраоптичного та паравентрикулярного ядер, що реагують на синаптичний приплив різної природи виділенням вазопресину та окситоцину; 4) нейрони ядер гіпоталамуса.
Дія основних класичних меадіаторів може бути відтворена за допомогою фармакологічних препаратів. Наприклад, нікотин викликає ефект, подібний до ефекту ацетилхоліну, при дії на постсинаптичну мембрану постгангліонарного нейрона, у той час як складні ефіри холіну і токсин мухомору мускарин - на постсинаптичну мембрану ефекторної клітини вісцерального органу. Отже, нікотин втручається у міжнейронну передачу в автономному ганглії, мускарин – у нейро-ефекторну передачу у виконавчому органі. На цій підставі вважають, що є відповідно два типи холінорецепторів: нікотинові (Н-холінорецептори) та мускаринові (М-холінорецептори). Залежно від чутливості до різних катехоламінів адренорецептори ділять на α-адренорецептори та β-адренорецептори. Їх існування встановлено за допомогою фармакологічних препаратів, що вибірково діють на певний вид адренорецепторів.
У ряді вісцеральних органів, що реагують на катехоламіни, є обидва види адренорецепторів, але результати їх збудження бувають, як правило, протилежними. Наприклад, у кровоносних судинах скелетних м'язів є α- та β-адренорецептори. Порушення α-адренорецепторів призводить до звуження, а β-адренорецепторів – до розширення артеріол. Обидва види адренорецепторів виявлені і в стінці кишки, проте реакція органу при збудженні кожного з видів однозначно характеризуватиметься гальмуванням активності гладких клітин м'язів. У серці та бронхах немає α-адренорецепторів і медіатор взаємодіє тільки з β-адренорецепторами, що супроводжується посиленням серцевих скорочень та розширенням бронхів. У зв'язку з тим, що норадреналін викликає найбільше збудження β-адренорецепторів серцевого м'яза і слабку реакцію бронхів, трахеї, судин, перші стали називати β1-адренорецепторами, другі - β2-адренорецепторами.
При дії на мембрану гладкої м'язової клітини адреналін і норадреналін активують аденілатциклазу, що знаходиться в клітинній мембрані. За наявності іонів Mg2+ цей фермент каталізує утворення у клітині цАМФ (циклічного 3",5"-аденозинмонофосфату) з АТФ. Останній продукт, у свою чергу, викликає ряд фізіологічних ефектів, активуючи енергетичний обмін, стимулюючи серцеву діяльність.
Особливістю адренергічного нейрона є те, що він має надзвичайно довгі тонкі аксони, які розгалужуються в органах і утворюють густі сплетення. Загальна довжина таких аксонних терміналей може досягати 30 см. По ходу терміналів є численні розширення - варикози, в яких синтезується, запасається та виділяється медіатор. З настанням імпульсу норадреналін одночасно виділяється з численних розширень, діючи відразу на велику площу гладком'язової тканини. Таким чином, деполяризація м'язових клітин супроводжується одночасним скороченням всього органу.
Різні лікарські засоби, що мають на ефекторний орган дію, аналогічну дії постгангліонарного волокна (симпатичного, парасимпатичного тощо), отримали назву міметиків (адрено-, холіноміметики). Поруч із є і речовини, вибірково блокують функцію рецепторів постсинаптичної мембрани. Вони названі гангліоблокаторами. Наприклад, амонієві сполуки вибірково вимикають Н-холінорецептори, а атропін та скополамін - М-холінорецептори.
Класичні медіатори виконують не тільки функцію передавачів збудження, але володіють і загальнобіологічною дією. До ацетилхоліну найбільш чутлива серцево-судинна система, він викликає і посилену моторику травного тракту, активуючи одночасно діяльність травних залоз, скорочує мускулатуру бронхів та знижує бронхіальну секрецію. Під впливом норадреналіну відбувається підвищення систолічного та діастолічного тиску без зміни серцевого ритму, посилюються серцеві скорочення, знижується секреція шлунка та кишки, розслаблюється гладка мускулатура кишки і т. д. Більш різноманітним діапазоном дій характеризується адреналін. За допомогою одночасної стимуляції іно-, хроно- та дромотропної функції адреналін підвищує серцевий викид. Адреналін надає розширюючу та антиспазматичну дію на мускулатуру бронхів, гальмує моторику травного тракту, розслаблює стінки органів, але гальмує діяльність сфінктерів, секрецію залоз травного тракту.
У тканинах всіх видів тварин виявлено серотонін (5-окситриптамін). У мозку він міститься переважно у структурах, що мають відношення до регуляції вісцеральних функцій, на периферії продукується ентерохромафінними клітинами кишки. Серотонін є одним з основних медіаторів метасимпатичної частини автономної нервової системи, що бере участь переважно в нейроефекторній передачі, і виконує також функцію медіаторію в центральних утвореннях. Відомо три типи серотонінергічних рецепторів - Д, М, Т. Рецептори Д-типу локалізовані в основному в гладких м'язах і блокуються діетіламідом лізергінової кислоти. Взаємодія серотоніну із цими рецепторами супроводжується м'язовим скороченням. Рецептори М-типу характерні більшості автономних гангліїв; блокуються морфіном. Зв'язуючись з цими рецепторами, передавач викликає гангліостимулюючий ефект. Рецептори Т-типу, виявлені в серцевій та легеневій рефлексогенних зонах, блокуються тіопендолом. Діючи на ці рецептори, серотонін бере участь у здійсненні коронарних та легеневих хеморефлексів. Серотонін здатний чинити пряму дію на гладку мускулатуру. У судинній системі воно проявляється у вигляді констрикторних чи дилататорних реакцій. При прямій дії скорочується мускулатура бронхів, при рефлекторному – змінюються дихальний ритм та легенева вентиляція. Особливо чутлива до серотоніну травна система. На введення серотоніну вона реагує початковою спастичною реакцією, що переходить в ритмічні скорочення з підвищеним тонусом і гальмуванням активності, що завершується.
Для багатьох вісцеральних органів характерною є пуринергічна передача, названа так унаслідок того, що при стимуляції пресинаптичних термінален виділяються аденозин та інозин – пуринові продукти розпаду. Медіатором у цьому випадку є А Т Ф. Місцем його локалізації служать пресинаптичні термінали ефекторних нейронів метасимпатичної частини автономної нервової системи.
АТФ, що виділився в синаптичну щілину, взаємодіє з пуринорецепторами постсинаптичної мембрани двох типів. Пуринорецептори першого типу чутливіші до аденозину, другого - до АТФ. Дія медіатора спрямовано переважно на гладку мускулатуру та проявляється у вигляді її релаксації. У механізмі кишкової пропульсії пуринергічні нейрони є головною антагоністичною гальмівною системою до збуджуючої холінергічної системи. Пуринергічні нейрони беруть участь у здійсненні низхідного гальмування, у механізмі рецептивної релаксин шлунка, розслаблення стравохідного та анального сфінктерів. Скорочення кишечника, що виникають за пуринергически викликаним розслабленням, забезпечують відповідний механізм проходження харчового грудки.
Серед медіаторів може бути гістамін. Він широко поширений у різних органах та тканинах, особливо у травному тракті, легенях, шкірі. Серед структур автономної нервової системи найбільша кількість гістаміну міститься у постгангліонарних симпатичних волокнах. На підставі реакцій у відповідь в деяких тканинах виявлені і специфічні гістамінові (Н-рецептори) рецептори: Н1- і Н2-рецептори. Класичною дією гістаміну є підвищення капілярної проникності та скорочення гладкої мускулатури. У вільному стані гістамін знижує кров'яний тиск, зменшує частоту серцевих скорочень, стимулює симпатичні ганглії.
На міжнейронну передачу збудження в гангліях автономної нервової системи гальмівний вплив має ГАМК. Як медіатор, вона може брати участь у виникненні пресинаптичного гальмування.
Великі концентрації різних пептидів, особливо субстанції Р, у тканинах травного тракту, гіпоталамуса, задніх корінців спинного мозку, а також ефекти стимуляції останніх та інші показники стали підставою вважати субстанцію Р медіатором чутливих нервових клітин.
Крім класичних медіаторів та «кандидатів» у медіатори, у регуляції діяльності виконавчих органів бере участь ще велика кількість біологічно активних речовин – місцевих гормонів. Вони регулюють тонус, коригують вплив на діяльність автономної нервової системи, їм належить істотна роль у координації нейрогуморальної передачі, в механізмах виділення та дії медіаторів.
У комплексі активних факторів чільне місце займають простагландини, яких багато міститься в волокнах блукаючого нерва. Звідси вони виділяються спонтанно чи під впливом стимуляції. Існує кілька класів простагландинів: Е, G, А, В. Їхня основна дія - збудження гладких м'язів, пригнічення шлункової секреції, релаксація мускулатури бронхів. На серцево-судинну систему вони мають різноспрямовану дію: простагландини класу А та Е викликають вазодилатацію та гіпотензію, класу G – вазоконстрикцію та гіпертензію.
Синапси ВНС мають загалом таку ж будову, що й центральні. Проте відзначається значне розмаїття постсинаптичних мембран хеморецепторів. Передача нервових імпульсів з прегангліонарних волокон на нейрони всіх вегетативних гангліїв здійснюється Н-холінергічні синапси, тобто. синапсами на постсинаптичній мембрані яких розташовані нікотинчутливі холінорецептори. Постгангліонарні холінергічні волокна утворюють на клітинах виконавчих органів (заліз, ГМК органів травлення, судин і т.д.) М-холінергічні синапси. Їхня постсинаптична мембрана містить мускаринчутливі рецептори (блокатор-атропін). І в тих та інших синапсах передача збудження здійснюється ацетилхоліном. М-холінергічні синапси мають збуджуючий вплив на гладкі м'язи травного каналу, сечовивідної системи (крім сфінктерів), залози ШКТ. Однак вони зменшують збудливість, провідність та скоротливість серцевого м'яза та викликають розслаблення деяких судин голови та тазу.
Постгангліонарні симпатичні волокна утворюють 2 типи адренергічних синапсів на ефекторах – a-адренергічні та b-адренергічні. Постсинаптична мембрана перших містить a1-і a2 – адренорецептори. При дії НА на a1-адренорецептори відбувається звуження артерій та артеріол внутрішніх органіві шкіри, скорочення м'язів матки, сфінктерів шлунково-кишкового тракту, але одночасно розслаблення інших гладких м'язів травного каналу. Постсинаптичні b-адренорецептори також поділяються на b1 – та b2 – типи. b1-адренорецептори розташовані у клітинах серцевого м'яза. При дії на них підвищується збудливість, провідність і скоротливість кардіоміоцитів. Активація b2-адренорецепторів призводить до розширення судин легень, серця та скелетних м'язів, розслаблення гладких м'язів бронхів, сечового міхура, гальмування моторики органів травлення
Крім того, виявлені постгангліонарні волокна, які утворюють на клітинах внутрішніх органів гістамінергічні, серотонінергічні, пуринергічні (АТФ) синапси.
2. вчення Павлова про 1 і 2 сигнальні системи.
Сигнальна система - система умовно- та безумовнорефлекторних зв'язків вищої нервової системи тварин (включаючи людину) та навколишнього світу. Розрізняють першу та другу сигнальні системи.
Термін запроваджено академіком І. П. Павловим.
Перша сигнальна система розвинена практично у всіх тварин, тоді як друга система присутня тільки у людини і, можливо, у деяких китоподібних. Це з тим, що людина здатна формувати абстрактний від обставин образ. Після вимовлення слова «лимон» людина може уявити, який він кислий і як зазвичай морщаться, коли їдять його, тобто вимовлення слова викликає у пам'яті образ (спрацьовує друга сигнальна система); якщо при цьому почалося підвищене відділення слини, це робота першої сигнальної системи.
Є предметом вивчення фізіології вищої нервової діяльності.
Друга сигнальна система - спеціальний тип вищої нервової діяльності людини, система "сигналів сигналів", що йдуть від загальної (але не однакової) з тваринами першої сигнальної системи - відчуттів, уявлень, що належать до навколишнього світу. Йдеться, як друга сигнальна система, як семіотична система значимостей - це «які у кору від мовних органів є другі сигнали, сигнали сигналів. Вони є відволікання від дійсності і допускають узагальнення, як і становить наше особисте, спеціально людське, вище мислення, що створює спочатку загальнолюдський емпіризм, а, нарешті, і науку - знаряддя вищої орієнтування людини у навколишньому світі й у собі». І. П. Павлов (1932).
Мозок тварини відповідає лише на безпосередні зорові, звукові та інші роздратування чи його сліди; Відчуття, що виникають, становлять першу сигнальну систему дійсності.
У процесі еволюції тваринного світу на етапі становлення та початкового розвитку виду Homo sapiens відбулася якісна видозміна системи сигналізації, що забезпечує активну та колективну адаптивну пристосувальну поведінку, яка створила різноманітні, прийняті в групі системи сигналізації та мови: слово, за висловом І. П. Павлова, стає "сигналом сигналів". Поява другої сигнальної системи - виникнення мови та мов, сигнальних систем людини з родичами, де умовні (довільні) сигнали індивіда набувають певних, прийнятих групою значення і значущості, перетворюються на знаки мови в прямому розумінні цього слова - це один із найважливіших результатів багатомільйоннолітньої еволюції соціальної життя роду Homo, що передаються через мовну діяльність з покоління до покоління.
У вивченні Ст с. с. спочатку переважало накопичення фактів, що характеризують значення узагальнюючої функції словесних сигналів, та був - розтин нервових механізмів дії слова. Встановлено, що узагальнення словом розвивається як результат вироблення системи умовних зв'язків (див. Умовні рефлекси); у своїй має значення як кількість зв'язків, а й їх характер: зв'язку, вироблені під час діяльності дитини, полегшують процес узагальнення. При дії словесних сигналів спостерігаються стійкі зміни збудливості, велика сила, частота і тривалість електричних розрядів у нервових клітинах певних пунктів кори мозку. Розвиток Ст с. с. - результат діяльності всієї кори великих півкуль; пов'язати цей процес із функцією якогось обмеженого відділу мозку неможливо. У дослідженнях В.С.С. у лабораторії вищої нейродинаміки та психології вищих когнітивних процесів Є. І. Бойка показано плідність вчення І. П. Павлова про динамічні тимчасові зв'язки В.с.с. У розвиток ідей І. П. Павлова та Є. А. Бойка у школі Є. А. Бойка розроблено загальну когнітивістську модель цілісного мовленнєво-мовного процесу, знайдено рішення найскладніших теоретичних проблем психології у її взаємозв'язках з лінгвістикою, такі як питання співвідношення мови і промови в процесах мовознавства та розуміння; характер зв'язків промови з думкою, промови з особистістю того, хто говорить; особливості розвитку дитячої мови та ін Тут розроблено нові методи аналізу публічних виступів (інтент-аналіз), що дозволяє певною мірою реконструювати «картину світу» мовця - його цільові та предметні спрямованості, їх динаміку, особливості в конфліктній ситуації, у вільних умовах спілкування, у громадських виступах та інших.
Істотним резервом для подальших досліджень залишаються проблеми типології колосальних індивідуальних відмінностей у взаємозв'язках загального та спеціального типів ВНД, неокортексу та емоційно-вольової та мимовільної регуляції діяльності та спілкування, поки що слабо представлених як у фізіології ВНД, так і в психолінгвістичних дослідженнях та в антропологічній ліні.
Подробиці
Гангліїявляють собою скупчення мультиполярних (один аксон і кілька дендритів) нейронів(Від кількох клітин до десятків тисяч). Екстраорганні (симпатичні) ганглії мають добре виражену сполучнотканинну капсулу, як продовження периневрію. Парасимпатичні ганглії перебувають, зазвичай, в интрамуральных нервових сплетіннях. Ганглії інтрамуральних сплетень, як і інші вегетативні вузли, містять вегетативні нейрони місцевих рефлекторних дуг. Мультиполярні нейрони діаметром 20-35 мкм розташовані дифузно, кожен нейрон оточений гліоцитами ганглію.
Крім того, описані нейроендокринні, хеморецепторні, біполярні, а у деяких хребетних та уніполярні нейрони. У симпатичних гангліях є дрібні інтенсивно флюоресцентні клітини (МІФ-клітини) з короткими відростками та великою кількістю гранулярних бульбашок у цитоплазмі. Вони виділяють катехоламіни і гальмують вплив на передачу імпульсів з прегангліонарних нервових волокон на еферентний симпатичний нейрон. Ці клітини називають інтернейрон.
Серед великих мультиполярних нейроніввегетативних гангліїврозрізняють: рухові (клітини Догеля 1-го типу), чутливі (клітини Догеля П-го типу) та асоціативні (клітини Догеля Ш-го типу). Двигуни нейрони мають короткі дендрити з пластинчастими розширеннями ("рецептивні майданчики"). Аксон цих клітин дуже довгий, йде за межі ганглію у складі постгангліонарних тонких безмієлінових нервових волокон і закінчується на гладких міоцитах внутрішніх органів. Клітини 1-го типу називають довгоаксонними нейронами. Нейрони П-го типу – рівновідросткові нервові клітини. Від їхнього тіла відходять 2-4 відростки, серед яких розрізнити аксон важко. Не розгалужуючись, відростки йдуть далеко від тіла нейрона. Їхні дендрити мають чутливі нервові закінчення, а аксон закінчується на тілах рухових нейронів у сусідніх гангліях. Клітини П-го типу є чутливими нейронами місцевих рефлекторних вегетативних дуг. Клітини Догеля Ш-го типу формою тіла схожі на вегетативні нейрони П-го типу, та їх дендрити не виходять межі ганглія, а нейрит прямує до інших ганглії. Багато дослідників вважають ці клітини різновидами чутливих нейронів.
Таким чином, у периферичних вегетативних ганглія є місцеві рефлекторні дуги, що складаються з чутливих, рухових і, можливо, асоціативних вегетативних нейронів.
Інтрамуральні вегетативні ганглії в стінці травного тракту відрізняються тим, що в їхньому складі, крім рухових холінергічних нейронів, є гальмівні нейрони. Вони представлені адренергічними та пуринергічними нервовими клітинами. Останніми медіатором є пуриновий нуклеотид. В інтрамуральних вегетативних гангліях зустрічаються також пептидергічні нейрони, що виділяють вазоінтестинальний пептид, соматостатин та ряд інших пептидів, за допомогою яких здійснюються нейроендокринна регуляція та модуляція діяльності тканин та органів травної системи.
Ацетилхолін- нікотинові (блок кураре, гексаметоній), мускаринові (блок атропін) рецептори. Активація рецепторів → генерування ВПСП. Швидкий ВПСП (N-холіноцер)→відкриття іонних каналів. Повільний ВПСП (М-холінорець)→пригнічення М-струму, обумовленого підвищенням К-провідності.
Нейропептиди- Діють як нейромодулятори.

Енкефаліни, Речовина Р, люліберин, нейротензин, соматостатин - симп. ганглії (+Ach)
Катехоламіни(NA, дофамін) – нейромедіатори дрібних клітин із інтенсивною флуоресценцією.
Нейропептид Y, соматостатин – симп. постгангліонари.
Симпатичні постгангліонари: NA, АТФ, нейропептид У.
α1→інозотолтрифосфат, діацилгліцерол. α2→активація G-білка, ↓цАМФ.
β→G-білок→AC→цАМФ
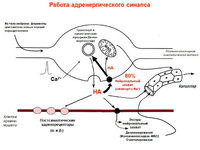
Винятки: медіатор Ach, мускаринові рецептори.
Парасимп. постгангліонари: Ach, ВІП, NO, соматостатин, АТФ, опіоїдні пептиди.
М1 (висока спорідненість до пірензепіну) - підвищення секреції кислоти клітинами залоз шлунка, М2 (низька) - уповільнюють серд. ритм, секреція слізних та слинних залоз.
Різноманітна дія:
-Специфічні втор. посередники: М2 може акт-ть IP3, і може инг-ть AC, зменшуючи цАМФ.
-Дія на К та Са-канали
-На ендотелії утворюється NO→гуанілатциклаза→цГМФ→цГМФ-залежна протеїнкіназа→розслаблення прасування.
У вегетативної нервової системирозрізняють центральні та периферичні відділи. Центральні відділи симпатичної нервової системи представлені ядрами бічних рогів тораколюмбального відділу спинного мозку. У парасимпатичній нервовій системі центральні відділи включають ядра середнього та довгастого мозку, а також ядра бічних рогів сакрального відділу спинного мозку. Парасимпатичні волокна краніобульбарного відділу виходять у складі III-й, VII-й, IX-й та Х-а парачерепні нерви.
Периферичні відділи вегетативної нервової системиутворені нервовими стовбурами, гангліями та сплетеннями.
Вегетативні рефлекторні дугипочинаються чутливим нейроном, тіло якого лежить у спинномозковому вузлі (ганглії), як і в соматичній рефлекторній дузі. Асоціативні нейрони перебувають у бічних рогах спинного мозку. Тут нервові імпульси перемикаються на проміжні прегангліонарні нейрони, відростки яких залишають центральні ядра і досягають вегетативних гангліїв, де передають імпульси на руховий нейрон. У зв'язку з цим розрізняють нервові волокна прегангліонарні та постгангліонарні. Перші з них залишають центральну нервову систему у складі вентральних корінців спинномозкових нервів та черепних нервів. Як у симпатичній, так і парасимпатичній системах преганглионарные нервові волокна належать холінергічним нейронам. Аксони нейронів, розташованих у вегетативних гангліях, називаються постгангліонарними. Вони не утворюють прямих контактів із ефекторними клітинами. Їх термінальні відділи за своїм ходом формують розширення - варикозності, у складі яких перебувають бульбашки медіатора. В області варикозності немає гліальної оболонки та нейромедіатор, виділяючись у навколишнє середовище, впливає на ефекторні клітини (наприклад, на клітини залоз, гладкі міоцити та ін.).
У периферичних гангліяхсимпатичної нервової системи, як правило, знаходяться адренергічні еферентні нейрони (за винятком нейронів, що мають синаптичні зв'язки з потовими залозами, де симпатичні нейрони є холінергічними). У парасимпатичних гангліях еферентні нейрони завжди холінергічні.
Гангліїє скупчення мультиполярних нейронів (від кількох клітин до десятків тисяч). Екстраорганні (симпатичні) ганглії мають добре виражену сполучнотканинну капсулу, як продовження периневрію. Парасимпатичні ганглії перебувають, зазвичай, в интрамуральных нервових сплетіннях. Ганглії інтрамуральних сплетень, як і інші вегетативні вузли, містять вегетативні нейрони місцевих рефлекторних дуг. Мультиполярні нейрони діаметром 20-35 мкм розташовані дифузно, кожен нейрон оточений гліоцитами ганглію. Крім того, описані нейроендокринні, хеморецепторні, біполярні, а у деяких хребетних та уніполярні нейрони. У симпатичних гангліях є дрібні інтенсивно флюоресцентні клітини (МІФ-клітини) з короткими відростками та великою кількістю гранулярних бульбашок у цитоплазмі. Вони виділяють катехоламіни і гальмують вплив на передачу імпульсів з прегангліонарних нервових волокон на еферентний симпатичний нейрон. Ці клітини називають інтернейрон.
Серед великих мультиполярних нейроніввегетативних гангліїв розрізняють: рухові (клітини Догеля І-го типу), чутливі (клітини Догеля ІІ-го типу) та асоціативні (клітини Догеля ІІІ-го типу). Двигуни нейрони мають короткі дендрити з пластинчастими розширеннями ("рецептивні майданчики"). Аксон цих клітин дуже довгий, йде за межі ганглію у складі постгангліонарних тонких безмієлінових нервових волокон і закінчується на гладких міоцитах внутрішніх органів. Клітини I типу називають довгоаксонними нейронами. Нейрони II типу - рівновідросткові нервові клітини. Від їхнього тіла відходять 2-4 відростки, серед яких розрізнити аксон важко. Не розгалужуючись, відростки йдуть далеко від тіла нейрона. Їхні дендрити мають чутливі нервові закінчення, а аксон закінчується на тілах рухових нейронів у сусідніх гангліях. Клітини II типу є чутливими нейронами місцевих вегетативних рефлекторних дуг. Клітини Догеля ІІІ-го типу за формою тіла схожі на вегетативні нейрони П-го типу, але їх дендрити не виходять за межі ганглію, а нейрит прямує в інші ганглії. Багато дослідників вважають ці клітини різновидами чутливих нейронів.
Таким чином, у периферичних вегетативних гангліяхє місцеві рефлекторні дуги, які з чутливих, рухових і, можливо, асоціативних вегетативних нейронів.
Інтрамуральні вегетативні гангліїу стінці травного тракту відрізняються тим, що в їхньому складі, крім рухових холінергічних нейронів, є гальмівні нейрони. Вони представлені адренергічними та пуринергічними нервовими клітинами. Останніми медіатором є пуриновий нуклеотид. В інтрамуральних вегетативних гангліях зустрічаються також пептидергічні нейрони, що виділяють вазоінтестинальний пептид, соматостатин та ряд інших пептидів, за допомогою яких здійснюються нейроендокринна регуляція та модуляція діяльності тканин та органів травної системи.
Навчальне відео анатомії вегетативної нервової системи (ВНС)
При проблемах із переглядом скачайте відео зі сторінкиАвтономні (вегетативні) нервові вузли (ганглії) можуть розташовуватися вздовж хребта (паравертебральні ганглії) або попереду нього (превертебральні ганглії), а також у стінці органів: серця, бронхів, травного тракту, сечового міхура та інших (інтрамуральні ганглії) або поблизу їх поверхні . Іноді вони мають вигляд дрібних (від кількох клітин до декількох десятків клітин) скупчень нейронів, розташованих по ходу деяких нервів або інтрамурально (мікроганглі). До вегетативних вузлів підходять прегангліонарні волокна (мієлінові), що містять відростки клітин, тіла яких лежать у центральній нервовій системі. Ці волокна сильно гілкуються та утворюють численні синаптичні закінчення на клітинах вегетативних вузлів. Завдяки цьому здійснюється конвергенція великої кількості терміналей прегангліонарних волокон на кожен нейрон ганглія. У зв'язку з наявністю синаптичної передачі вегетативні вузли відносять до нервових центрів ядерного типу.
Вегетативні нервові вузли за функціональною ознакою та локалізації поділяються на:
симпатичні;
парасимпатичні.
Симпатичні нервові вузли(пара-і превертебральні) отримують прегангліонарні волокна від клітин, розташованих у вегетативних ядрах грудних та поперекових сегментів спинного мозку. Нейромедіатором прегангліонарних волокон є ацетилхолін, а постгангліонарних – норадреналін (за винятком потових залоз та деяких кровоносних судин, що мають холінергічну симпатичну іннервацію). Крім цих нейромедіаторів, у вузлах виявляються енкефаліни, речовина Р, соматостатин, холецистокінін.
Парасимпатичні нервові вузли(інтрамуральні, що лежать поблизу органів або вузли голови) отримують прегангліонарні волокна від клітин, розташованих у вегетативних ядрах довгастого та середнього мозку, а також крижового відділу спинного мозку. Ці волокна залишають центральну нервову систему у складі 3, 7, 9, 10 пар черепномозкових нервів та передніх корінців крижових сегментів спинного мозку. Нейромедіатором пре-і постгангліонарних волокон є ацетилхолін. Крім нього роль медіаторів у цих гангліях грають серотонін, АТФ, можливо, деякі пептиди.
Більшість внутрішніх органів має подвійну вегетативну іннервацію, тобто одержують постгангліонарні волокна від клітин, розташованих як у симпатичних, так і парасимпатичних вузлах. Реакції, що опосередковуються клітинами симпатичних та парасимпатичних вузлів, часто мають протилежну спрямованість, наприклад: симпатична стимуляція посилює, а парасимпатична – гальмує серцеву діяльність.
Загальний план будови симпатичних та парасимпатичних нервових вузлів подібний. Вегетативний вузол покритий сполучнотканинною капсулою і містить дифузно або групами розташовані тіла мультиполярних нейронів, їх відростки у вигляді безмієлінових або, рідше, мієлінових волокон та ендоневрій. Тіла нейронів мають неправильну форму, містять ексцентрично розташоване ядро, оточені (зазвичай не повністю) оболонками із гліальних клітин-сателітів (мантійних гліоцитів). Часто зустрічаються багатоядерні та поліплоїдні нейрони.
Інтрамуральні вузли та пов'язані з ними провідні шляхи через їх високу автономію, складність організації та особливості медіаторного обміну деякими авторами виділяються в самостійний метасимпатичний відділ вегетативної нервової системи. Зокрема, загальна кількість нейронів в інтрамуральних вузлах кишки вища, ніж у спинному мозку, а за складністю їхньої взаємодії в регуляції перистальтики та секреції їх порівнюють з мінікомп'ютером.
В інтрамуральних вузлах описані нейрони трьох типів:
довгоаксонні еферентні нейрони (клітини Догеля І типу) чисельно переважають. Це великі або середніх розмірів еферентні нейрони з короткими дендритами та довгим аксоном, що прямує за межі до робочого органу, на клітинах якого він утворює рухові або секреторні закінчення;
рівновідросткові аферентні нейрони (клітини Догеля II типу) містять довгі дендрити і аксон, що виходить за межі даного ганглія в сусідні і утворює синапси на клітинах І і ІІІ типів. Ці клітини, мабуть, входять як рецепторна ланка до складу місцевих рефлекторних дуг, які замикаються без заходу нервового імпульсу до центральної нервової системи. Наявність таких дуг підтверджується збереженням функціонально активних аферентних, асоціативних та еферентних нейронів у трансплантованих органах (наприклад, серце);
асоціативні клітини (клітини Догеля ІІІ типу) — місцеві вставні нейрони, що з'єднують своїми відростками кілька клітин І та ІІ типу, морфологічно подібні з клітинами Догеля ІІ типу. Дендрити цих клітин не виходять за межі вузла, а аксони прямують до інших вузлів, утворюючи синапси на клітинах I типу.


